
泛素化,顾名思义,肯定需要泛素的参与。泛素-蛋白酶体系统 (UPS),则同时与泛素和蛋白酶体有关。
真核细胞中的两种主要降解系统:泛素-蛋白酶体系统和溶酶体降解系统。在真核细胞内,80% 以上的蛋白质的降解都依赖于“泛素-蛋白酶体系统”。
这一降解过程可以分为两个阶段:(1) 给废弃蛋白质“贴标签”: 泛素与底物蛋白的相互作用,底物带上泛素标签;(2) “标签”蛋白质进行“废物处理”: 经过泛素化修饰的蛋白质被特定的蛋白酶体识别,从而被降解[1]。
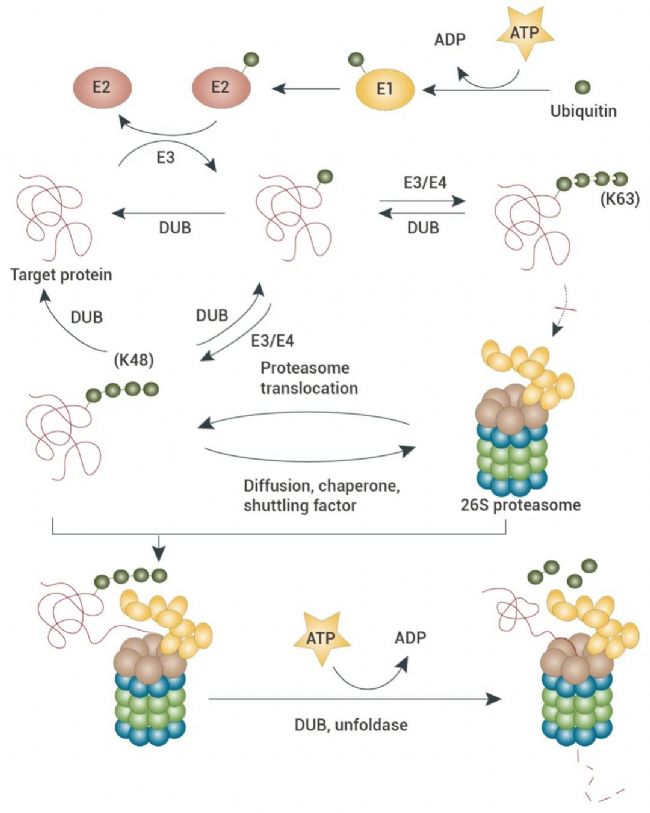
图 1. 泛素介导的蛋白质降解过程[1]。
首先,泛素与 E1 (泛素激活酶) 共价结合,然后被转移至 E2 (泛素结合酶) 上。接着,E3 (泛素-蛋白质连接酶) 将泛素从 E2 转移到底物蛋白上,并在 E4 酶 (链延长因子) 的催化下延长链长,最终使底物蛋白带上了泛素标签。相关调节颗粒可以识别连接在蛋白质上的多泛素链标签,并启动降解过程[1]。
蛋白酶体除了通过上述经典的“泛素-蛋白酶体系统”降解蛋白以外,还可以通过非泛素化修饰降解蛋白。例如,已有研究表明,Fos 家族可通过泛素化依赖的和不依赖的两种机制被降解[2]。但一直以来,关于 IEG 蛋白 (Immediate-early genes) 迅速靶向降解的机制,尚不清楚,直到……
 近日,来自哈佛医学院的 Michael E. Greenberg 教授团队在顶尖期刊 Science 上发布了一项突破性的研究成果。
近日,来自哈佛医学院的 Michael E. Greenberg 教授团队在顶尖期刊 Science 上发布了一项突破性的研究成果。
Michael E. Greenberg 团队通过全基因组 CRISPR-Cas9 筛选调控 IEGs 蛋白稳定性的基因,发现了一种哺乳动物中基本未被表征过的 Midnolin 蛋白,其可以促进不同 IEGs 蛋白的蛋白酶体降解。进而发现了一种全新的蛋白降解机制——Midnolin-蛋白酶体途径。
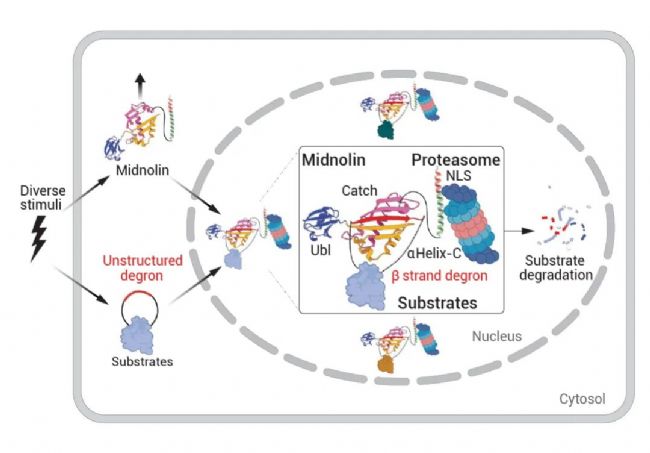
图 2. Midnolin-蛋白酶体途径降解核蛋白过程[1]。
Midnolin 受多种因素诱导,包括生长因子和神经元刺激。在细胞核内,Midnolin 通过 C 端 α 螺旋 (αHelix-C) 与蛋白酶体结合,利用 Catch 结构域实现选择性,并通过 N 端泛素样结构域 (Ubl) 促进结合底物的降解 (图 2)。研究发现,Midnolin-蛋白酶体途径绕过典型的泛素化系统来实现许多核蛋白的选择性降解[2]。
 早期研究:
早期研究:Midnolin 最初由 T sukahara 等人在胚胎干细胞中发现 (2000 年)[4]。
(1) 利用基因捕获方法 (Gene trap) 和体外胚胎干细胞的选择,确定了参与核仁神经发生相关基因调控的新基因--Midnolin。
(2) Midnolin 编码一种含有 508 个氨基酸 (aa) 的蛋白质,其中包含一个泛素样结构域。
(3) 利用 Midnolin-绿色荧光蛋白 (GFP) 融合蛋白研究 Midnolin 在细胞内的分布 。Midnolin 被发现定位于细胞核和核仁,但不在细胞质中。核仁定位信号 (NLS) 为位于 Midnolin C 端区域的包含 28 个氨基酸的肽 (440-QQKRLRRKARRDARGPYHWTPSRKAGRS-467)。
▐ Midnolin 是 IEG 蛋白质降解的调节剂
为了研究 IEGs 蛋白质降解的机制,作者重点研究了两种常见的来自不同蛋白家族的转录因子——EGR1 和 Fos。通过构建稳定表达 EGR1 或 FosB GPS (The global protein stability) 报告基因的 HEK-293T 细胞系,并进行了全基因组 CRISPR-Cas9 筛选,发现 MIDN 是负调节 EGR1 和 FosB 稳定性的热点基因。并发现敲除 MIDN 基因会增加 EGR1 和 FosB 稳定性。
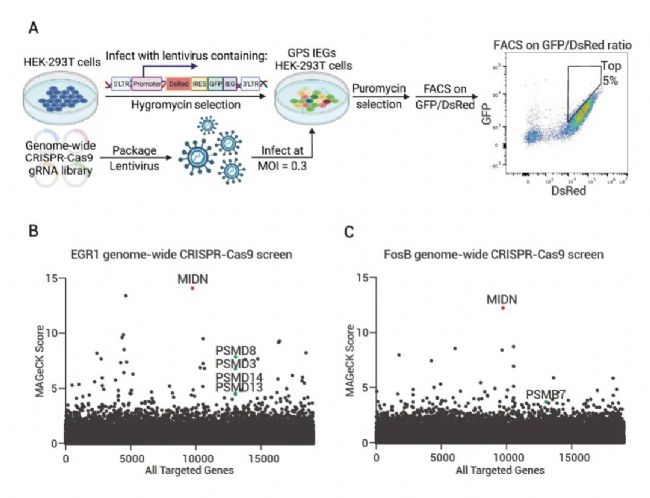
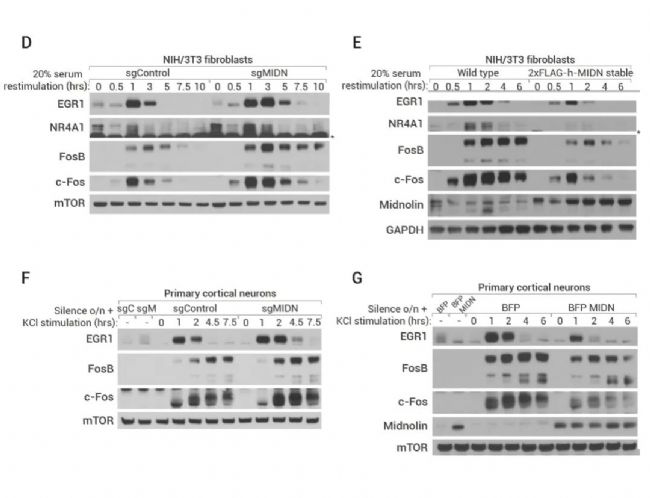
图 3. Midnolin 是 IEGs 蛋白降解的调节剂,可促进多种 IEG 蛋白的降解[2]。
A-C:遗传学筛选显示 Midnolin 是 IEGs 蛋白降解的调节。(A) 构建稳定表达 EGR1 或 FosB GPS 报告基因的 HEK-293T 细胞系并进行全基因组 CRISPR-Cas9 筛选示意图。(B-C) 遗传筛选结果,显示 MIDN 是负调节 EGR1 和 FosB 稳定性的热点基因。D-G:Midnolin促进生理环境中多种 IEG 蛋白的降解。(D-E) NIH/3T3 细胞中敲除/过表达 Midnolin 对 EGR1、FosB、c-Fos 和 NR4A1 蛋白水平的影响; (F-G) 初级皮层神经元中敲除/过表达 Midnolin 对 EGR1、FosB、c-Fos 和 NR4A1 蛋白水平的影响。
此外,在 NIH/3T3 成纤维细胞 (图 3D-E) 和初级皮层神经元 (图 3F-G) 中过表达 Midnolin 降低了多种 IGEs 蛋白水平,如 EGR1、FosB、c-Fos 和 NR4A1,且敲除 Midnolin 结果与之相反。上述结果表明这些 IEGs 蛋白可能都通过 Midnolin 发生靶向降解。进一步研究显示,Midnolin 可能还参与了细胞核内除 Fos 和 EGR1 外数百种其他转录因子的降解。
作者将内源性 Midnolin 用 3xHA 标记,进行质谱分析发现 Midnolin 可免疫共沉淀 19 S 调节颗粒和 20 S 核心颗粒体 (图 4A)。同时,使用 MG132 (蛋白酶体抑制剂) 抑制底物蛋白的降解,发现 Midnolin 与底物 c-Fos、FosB、EGR1 存在相互作用,且与 PSMD2 和 PSMA2 (分别为 19 S 和 20 S 蛋白酶体的组成部分) 存在相互作用 (图 4B)。
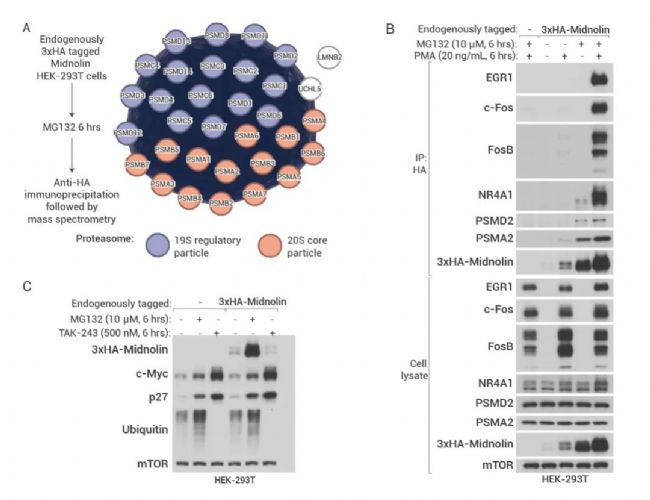
(A) 将 3xHA 标签引入 HEK-293T 细胞中内源性 Midnolin 的 N 末端。细胞用 MG132 处理 6 小时后进行 3xHA-Midnolin 免疫沉淀,并进行质谱分析。(B) Midnolin 免疫共沉淀蛋白酶体和 内源 IEGs 蛋白。PMA: PKC 激动剂,诱导 IEGs 的转录。(C) 内源性 Midnolin 水平因抑制蛋白酶体而显着增加,但不会因抑制泛素 E1 而增加。
此外,MG132 可增加 Midnolin 蛋白水平,但 TAK-243 (泛素 E1 酶抑制剂) 则不影响 Midnolin 蛋白水平 (图 4C),且 TAK-243 并不影响 Midnolin 对底物蛋白的降解,由此证明 Midnolin 降解蛋白不依赖于泛素化。
注:26S 蛋白酶体由中间一个圆柱形 20S 核心颗粒和两端覆盖的一个或两个 19S 调节颗粒组成。
▐ Midnolin 含三个协同作用的结构域,促进结合底物的蛋白酶体降解
作者通过使用人工智能平台 AlphaFold 进行结构预测,显示 Midnolin 蛋白包含三个稳定的结构域: 位于 N 末端的泛素样结构域 (Ubl)、含两部分假对称 (pseudosymmetric) 的“捕获 Catch”结构域、和一个位于 C 末端的长 α 螺旋 (α Helix-C) (图 5A)。
接下来,作者使用免疫共沉淀实验以确定 Midnolin 与底物和/或蛋白酶体相互作用所需的结构域。结果显示,Ubl 结构域的点突变/缺失/诱变对 Midnolin 与EGR1 或蛋白酶体的相互作用无影响,但 Catch 结构域的缺失使得底物蛋白 (如 EGR1) 无法被沉淀下来 (图 5C 左),α Helix-C 或 NLS 结构域的缺失使得蛋白酶体 (如 PSMD2) 无法被捕获 (图 5C 右)。
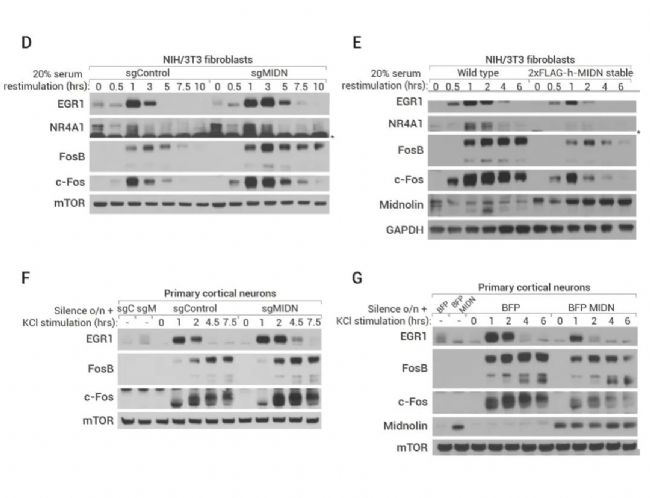
图 5. Midnolin 包含三个结构域,它们协同作用以促进底物的蛋白酶体降解[2]。
(A) AlphaFold 预测的 Midnolin 结构; (B) 引入 Midnolin cDNA 的突变或截短的示意图; (C) 使用 CMV 启动子对 HEK-293T 细胞稳定表达 2XFlag Midnolin,用 10 μM MG132 处理细胞 6 小时,并进行免疫印迹。
也就是说,Midnolin 通过 Catch 结构域与目标蛋白相互作用,通过 α Helix-C 结构域与 26S 蛋白酶体结合。同时,Midnolin 可通过其 Catch 结构域结合底物内的非结构化区域 (unstructured regions),并形成 β 链。虽然 Midnolin-蛋白酶体途径可绕过泛素化系统来实现许多核蛋白的选择性降解,但关于如何实现特异性的靶蛋白降解还有待深入研究。

本期小 M 带大家回顾了经典的泛素化蛋白质降解过程,并介绍了一种全新蛋白降解机制 —— Midnolin-蛋白酶体途径,该途径可绕过泛素化系统来实现许多核蛋白的选择性降解。小 M 相信,基于 Midnolin 的新型靶向蛋白降解类药物将会被逐渐开发出来,用于神经或精神疾病,癌症等疾病的研究~

|
蛋白酶体抑制剂 (IC50: 100 nM),可诱导凋亡和自噬。 |
|
泛素 E1 酶 (UBA1) 抑制剂,抑制蛋白泛素化。 |
|
PKC 和 SphK 激活剂,可诱导 IEGs 的转录。 |
|
200+ 活性化合物,用于泛素化及相关疾病研究。 |
MCE的所有产品仅用作科学研究或药证申报,我们不为任何个人用途提供产品和服务。
参考文献:
[1] Tai HC, et al. Ubiquitin, the proteasome and protein degradation in neuronal function and dysfunction. Nat Rev Neurosci. 2008 Nov;9(11):826-38.
[2] Gu X, et al. The midnolin-proteasome pathway catches proteins for ubiquitination-independent degradation. Science. 2023 Aug 25;381(6660):eadh5021.
[3] Obara Y, et al. Midnolin is a novel regulator of parkin expression and is associated with Parkinson's Disease. Sci Rep. 2017 Jul 19;7(1):5885.
[4] Tsukahara M, et al. Novel nucleolar protein, midnolin, is expressed in the mesencephalon during mouse development. Gene. 2000 Aug 22;254(1-2):45-55.